Monthly Archives: gener 2024
El cervell humà es va desenvolupar gràcies als aliments fermentats
19 gener 2024
Click here for the English version: Human brain developed thanks to fermented food
Estic subscrit, entre altres mitjans digitals, a Science Alert, un newsletter —butlletí informatiu— científic gratuït, en el qual cada setmana trobo alguna notícia interessant. Fa poques setmanes em va sorprendre molt especialment una notícia (Dyer R (2023) sobre aquesta hipòtesi del títol, la qual trobo molt versemblant. A banda d’aquesta notícia-resum, per conèixer-ne millor els detalls vaig anar a l’article original de Bryant et al. “Fermentation technology as a driver of human brain expansion”, que us comento a continuació, junt amb altres coses relacionades.
CERVELL MÉS GRAN
La principal característica diferencial dels humans respecte als altres primats i animals en general, és el cervell més gran i complex. Com que quan més gran és un animal, major és el pes de l’encèfal, s’utilitza una mesura relativa que és el quocient d’encefalització (EQ), que és la relació entre la massa de l’encèfal i l’esperada per a un animal típic de les mateixes dimensions. L’EQ de Homo sapiens és al voltant de 7.5, mentre que per a altres primats és entre 2 i 3, i altres mamífers com el gos és entre 1 i 2, excepte els cetacis com orques o dofins, que tenen 3 o 4.
Per tant, el cervell humà va més que triplicar la seva grandària respecte a altres primats en la seva evolució des dels darrers australopitecs —que ja eren bípedes, i per tant tenien les mans lliures— fa uns 2.5 milions d’anys (Ma) fins als primers Homo aproximadament mig milió d’anys després. Els australopitecs tenien un volum endocranial (ECV) de 400 mL (Figura 1), mentre que per a Homo erectus (Figura 2) era d’uns 800 mL, i després l’expansió del cervell va continuar amb l’emergència de H. sapiens i prèviament amb H. neanderthalensis, ambdós amb uns 1500 mL (Miller et al. 2019; Ponce de León et al. 2021). Òbviament aquest engrandiment del cervell i sobretot del còrtex frontal va determinar l’augment de les capacitats de raonament, reflexió, adaptació, socialització i d’altres habilitats, o sigui del desenvolupament de la intel·ligència humana.


MÉS CERVELL i MENYS BUDELLS
Hi ha diverses teories sobre els mecanismes que haurien afavorit aquesta accelerada expansió del cervell. El factor limitant en l’engrandiment és la disponibilitat de recursos calòrics, perquè el cervell té una alta despesa metabòlica, comparada amb la majoria d’altres teixits. La taxa metabòlica en repòs del cervell suposa el 22% de la del cos humà (McClave & Snider 2001).
Les mutacions que van portar a un augment de mida del cervell, encara que tindrien uns clars beneficis finals, no serien adaptatives si comportaven un major risc de fam. Una reducció en la quantitat de teixit intestinal, que té unes necessitats metabòliques similars a les del cervell, alliberaria calories necessàries per a la digestió per tal de reassignar-les al cervell. Això es confirma pel fet que la mida del còlon dels humans és la quarta part del corresponent a primats de la nostra talla (Taula 1), mentre que el cervell dels H. sapiens actuals és quasi el triple del que correspondria.
Taula 1. Masses esperades d’alguns òrgans humans basades en els valors dels grans simis en comparació amb les reals d’un humà occidental actual de 65 kg (Adaptat de Bryant et al 2023).
| Òrgan | Massa esperada (kg) | Massa real (kg) | Real / esperada |
| Cor | 0.32 | 0.30 | 0.94 |
| Fetge | 0.24 | 0.30 | 1.25 |
| Budell prim | 0.40 | 0.62 | 1.55 |
| Còlon | 0.85 | 0.22 | 0.26 |
| Cervell | 0.45 | 1.30 | 2.89 |
CANVIS DE DIETA
Tanmateix, la reducció intestinal havia d’anar acompanyada d’un canvi a la dieta, amb aliments més fàcils de digerir i més energètics. Els homínids precursors d’Homo haurien passat d’un règim herbívor-frugívor a un omnívor-carnívor. Les hipòtesis actuals apunten als dos canvis següents, força coneguts i bastant versemblants:
1) El major consum de carn —de proteïna animal en general— s’ha argumentat com un dels elements clau en l’evolució humana. La dieta de H. sapiens és clarament més carnívora que els altres primats parents, i per tant la cacera d’altres animals hauria d’haver estat un hàbit creixent als precursors de Homo (Mann 2000). Tanmateix, un punt feble d’aquesta hipòtesi és que sembla que la caça era poc important inicialment, fa 1-2 Ma, ja que aquests primers Homo i els seus precursors eren sobretot recol·lectors, i la cacera es desenvolupà plenament força després, a finals del paleolític inferior —com a molt fa uns 500.000 anys—, en paral·lel al desenvolupament de les primeres armes prehistòriques (Bryant et al. 2023).
En canvi, sembla que el consum de carronya deixada per altres animals carnívors fou anterior a la cacera, des del final del pliocè o inicis del plistocè fa uns 3 Ma. Una alternativa relacionada amb la necrofàgia però més profitosa per consumir carn és prendre les preses caçades per altres animals. De fet, els registres arqueològics demostren que aquesta darrera opció per obtenir carn fresca era predominant sobre la de ser carronyaires passius, on el rendiment de consum càrnic és més baix (Bunn & Ezzo 1993).
Però el consum de proteïna animal d’alt valor nutritiu no es limita a la carn de mamífers i aus: cal tenir en compte la pesca i sobretot la recol·lecció de marisc. Aquest darrer cas té l’avantatge arqueològic d’haver-se trobat molts llocs amb closquers —acumulació de closques de mol·luscs— que indiquen la gran explotació d’aquest recurs proteic que els humans han fet de fa temps. Els closquers trobats més antics són els de Pinnacle Point a Sudàfrica fa 160.000 anys, molt importants perquè junt amb altres restes són una de les evidències dels primers Homo sapiens (Marean et al. 2007), però és clar, són certament posteriors al desenvolupament del cervell.
2) La domesticació del foc i la conseqüent possibilitat de cuinar els aliments va ser un altre element crucial per obtenir més substrats calòrics biodisponibles i digerir-los més fàcilment, tant a nivell de masticació mecànica com de despesa energètica necessària al tracte digestiu. Això és molt evident en el consum de carn comentat, tant fresca com de carronya, i a més en aquest cas per mitigar la contaminació microbiana. A més a més el cuinat també va ser molt important en permetre la ingestió d’aliments vegetals i sobretot dels tubercles, òrgans de reserva soterranis molt rics en carbohidrats, però que no són directament digeribles i/o contenen compostos tòxics si no es couen (Wrangham et al. 1999).
Tanmateix, no hi ha evidències arqueològiques clares de què els australopitecs o els primers Homo dominessin el foc, i les primeres evidències serien per als H. erectus entre 1 i 1.5 Ma (Hlubik et al. 2019) però més clarament de fa 800.000 anys (Goren-Inbar et al 2004). Per tant, el ple domini del foc hauria estat posterior a l’inici del desenvolupament del cervell. De fet, l’expertesa del foc requereix la capacitat cognitiva de planejar, crear, mantenir i usar el foc efectivament, o sigui, un cervell més desenvolupat que el dels australopitecs (Bryant et al. 2023).
HIPÒTESI DE LA FERMENTACIÓ EXTERNA dels aliments, en contrast amb l’habitual “INTERNA”
Els autors del treball (Bryant et al. 2023) proposen aquest terme d’externa per diferenciar-lo de la fermentació interna que és la realitzada per la microbiota del tracte gastrointestinal humà en la digestió. La idea és que l’externalització d’aquesta fermentació interna va alliberar requeriments energètics corporals que van permetre l’expansió cerebral.
Cal recordar que aquí s’utilitza el terme “fermentació” en la seva accepció més general de transformació de compostos orgànics per part de microorganismes, mentre que l’accepció original del concepte “fermentació” en sentit bioquímic estrictament és el tipus de metabolisme heteròtrof anaeròbic on font d’energia i carboni són compostos orgànics i l’acceptor d’electrons són aquests mateixos compostos. La majoria de fermentacions d’aliments com la làctica o alcohòlica són d’aquesta accepció bioquímicament parlant, però altres processos de transformació microbiana que incloem a més d’aquestes quan es parla de “fermentació” en general, són d’altres tipus de metabolisme, com per exemple degradacions aeròbiques o altres reaccions. Podeu veure més informació sobre els aliments fermentats al meu post “Aliments fermentats: consens recent sobre aquest concepte” (Figura 3).

Encara que no és usual anomenar-ho així, la digestió que té lloc al tracte gastrointestinal humà o d’altres animals inclou aquesta “fermentació interna”, entenent com a tal la intervenció microbiana, o sigui el conjunt de transformacions que hi realitzen els microorganismes, la microbiota intestinal, sobretot al còlon. La digestió d’una part important dels components fibrosos vegetals requereix aquesta fermentació interna per part de la microbiota. Als remugants això s’aconsegueix a més amb estómacs addicionals i una microbiota cel·lulolítica abundant. Als altres animals no remugants, inclosos els primats, tenim un còlon i un cec més desenvolupats, i una major àrea per a l’absorció dels nutrients. El còlon dels humans i molts primats conté uns 10^12 microbis per mL i el trànsit per aquest budell gros és d’unes 20-40 hores, mentre que al prim només és 2-4 h. Cada cop és més clara la rellevància de la microbiota del còlon per a la salut, en quant a l’absorció de nutrients, la regulació energètica i un sistema immune correcte (O’Hara & Shanahan 2006).
La fibra soluble, sobretot oligosacàrids, és fermentada per la microbiota produint sobretot àcids grassos de cadena curta (SCFA), que aporten unes 2 cal/g de fibra, que suposa un 50% addicional a les 4 cal/g disponibles per la digestió directa dels carbohidrats fàcils (midó, sucres). Aquestes 2 cal/g són un 2-10% de l’energia total que la dieta ens proporciona, que és molt poca comparada amb altres mamífers (Taula 2). Aquests SCFA són sobretot acetat, propionat i butirat. L’acetat s’utilitza per sintetitzar colesterol i altres SCFA i proporciona energia al cor, ronyons i múscul; el propionat és precursor de la síntesi hepàtica de glucosa i proteïnes; i el butirat és la font d’energia preferida dels colonòcits.
Taula 2. Energia derivada dels àcids grassos de cadena curta (SCFA) produïts per la fermentació de la microbiota intestinal
| Espècie | Dieta | % Energia del total digerit |
| Bou / vaca | Herbívor remugant | 72 |
| Ovella | Herbívor remugant | 84 |
| Conill | Herbívor monogàstric | 32 |
| Castor | Herbívor monogàstric | 19 |
| Porc espí | Herbívor monogàstric | 16 |
| Porc | Omnívor | 36 |
| Aluata, mico udolador | Herbívor monogàstric | 30 |
| Goril·la | Herbívor monogàstric | 57 |
| Homo sapiens | Omnívor | 2-10 |
A banda dels SCFA, els nutrients principals produïts per la microbiota són les vitamines del complex B i la K, que són absorbides per l’intestí. A més, la microbiota augmenta la biodisponibilitat de micronutrients minerals mitjançant la degradació de factors antinutricionals com els fitats i oxalats —presents a molts vegetals— que formen complexos amb els cations (Fe, Zn, Mg, Ca, …) i prevenen la seva absorció.
La fermentació externa dels aliments que van començar a fer els primers humans té unes funcions similars a la interna com l’augment de la biodisponibilitat i absorció de macronutrients i micronutrients. Amb això, s’augmenta la digestibilitat de carbohidrats i proteïnes, per exemple als llegums hidrolitzant les macromolècules a aminoàcids i sucres més digeribles. La fermentació de carbohidrats també fa augmentar les vitamines B disponibles en un ordre de magnitud (Sandhu et al 2017). I els comentats fitats i oxalats poden ser degradats per la fitasa que produeixen bacteris làctics de les fermentacions externes, amb la qual cosa augmenta l’absorció de minerals. Aquesta eliminació del fitat fins i tot és més efectiva fermentant que per cocció, ja que amb aquesta baixa l’activitat de la fitasa per damunt de 80ºC.
Un gran benefici de la fermentació externa és que pot fer que aliments tòxics ho deixin de ser. El cas més conegut és la destoxificació dels glicòsids cianogènics de la mandioca (o iuca o cassava), un aliment bàsic de milions de persones a les zones tropicals. Si no es fermenta, aquests glicòsids són hidrolitzats pels microbis del còlon produint el tòxic cianur. Quan es fermenta adientment els bacteris làctics trenquen les parets cel·lulars dels tubercles i permeten la hidròlisi de la toxina, afavorida també per l’àcid làctic que produeixen (Padmaja & Steinkraus 1995).
A més a més, la fermentació externa dels aliments contribueix a una millor eficàcia de la microbiota intestinal en la digestió. En primer lloc, part de la microbiota ingerida amb l’aliment fermentat pot colonitzar l’intestí, contribuint a la seva biodiversitat, augmentant la capacitat de fermentar més nutrients, i afavorint que alguns microbis endògens produeixin bacteriocines contra possibles patògens. S’ha vist que aquests beneficis també són possibles encara que els microbis de l’aliment fermentat només tinguin un contacte transitori amb els bacteris residents (Ohland & MacNaughton 2010). Amb això, la fermentació externa pot ajudar a la microbiota endògena a protegir l’hoste d’infeccions i malalties, ja que una microbiota correcta produint quantitat de SCFA a partir de la fermentació de carbohidrats no digeribles està ben relacionada amb una reducció dels desordres gastrointestinals (Alexander et al 2019).
FERMENTACIÓ EXTERNA d’ALIMENTS, IMPULSORA DE L’EXPANSIÓ DEL CERVELL HUMÀ
Com hem vist abans, sembla que els canvis de dieta des dels australopitecs als humans, com el major consum de carn o d’aliments cuinats amb el domini del foc, són relativament posteriors a l’expansió del cervell, i només amb aquests canvis no s’acaba d’explicar el ràpid desenvolupament del cervell, simultània a la reducció del còlon i el desplaçament de gran despesa energètica de l’intestí al cervell.
Per als inicis de la fermentació externa d’aliments no caldria tenir una gran capacitat de raonament. Els australopitecs ja tenien algunes eines senzilles que podrien utilitzar per escorxar animals capturats o de la carronya, i podrien transportar aquests aliments fins a l’habitatge, fos cova o balma o refugis temporals, gràcies al bipedisme ja desenvolupat. També podrien transportar fruits, tubercles i altres aliments potencials. Encara que per exemple els ximpanzés ocasionalment poden transportar eines temporals o les restes d’animals caçats, ho fan en distàncies curtes, d’uns centenars de metres com a molt, i la majoria dels aliments els consumeixen al lloc de captura.
Un cop al lloc d’habitatge, aquests primers Homo devien deixar el menjar per anar-lo consumint i acumulant-ne més del capturat. La reutilització de lloc d’emmagatzematge hauria promogut un ecosistema microbià que conduís a la fermentació. Els aliments novament incorporats haurien estat inoculats amb els ja presents al lloc, o al cos dels mateixos homínids, les mans per exemple. Aquesta pràctica transmesa socialment de reutilitzar llocs, contenidors o eines per manipular els aliments hauria anat promovent les fermentacions i l’estabilitat dels agents microbians fermentatius. Com en tot procés de selecció, aquesta tecnologia primitiva s’hauria anat modificant, sobretot aprenent a no consumir els productes fets malbé amb patògens o compostos tòxics, segurament amb més d’una víctima pel camí.
La fermentació externa dels aliments requereix pocs coneixements, força menys que la utilització del foc, ja que la fermentació és un procés natural que pot passar espontàniament, és un procés passiu per al qual no cal un esforç actiu com el de mantenir el foc. I a més, la fermentació pot preservar els aliments durant molt de temps, anys i tot, gràcies sobretot a alguns productes de la fermentació com l’àcid làctic o l’etanol.
De fet, es proposa que altres espècies de Homo com els Neandertals ja deixaven fermentar carn, amb la qual cosa la baixada de pH degut a l’àcid produït preservava la vitamina C continguda a la carn i així evitaven l’escorbut (Speth 2019).
Segurament la fermentació s’aniria combinant amb altres tècniques de conservació com el fumat, assecat i salat, com es fa actualment. Però la facilitat de la fermentació en molt diversos tipus d’aliments, ambients i condicions devia permetre la seva difusió. La prova més evident és que en l’actualitat hi ha múltiples aliments fermentats, a pràcticament totes les parts del món. Es calcula que hi ha més de 5000 varietats d’aliments fermentats, que segons la FAO són el 35% del mercat actual de tots els aliments. En veiem alguns a la Taula 3 i a la Figura 4. També podem veure una selecció de 36 d’ells a un lloc web d’informació gastronòmica: howtocook.recipes.
Taula 3. Relació d’aliments fermentats, ordenats pels tipus de substrat: parts de vegetals o animals (modificada i ampliada de Bryant et al 2023).
| Nom producte | Substrat | Tipus producte | Lloc d’origen | Microorganismes |
| Kimchi | fulles de col, rave i altres | àcid | Àsia E | Bacteris làctics (BL) |
| Xucrut | fulles de col | àcid | Europa | BL, enterobacteris |
| Pu-erh | fulles de té | àcid, beguda | Àsia E | Floridures, llevats |
| Kombutxa de té | fulles de té | àcid, beguda | Àsia E | Bacteris acètics, llevats |
| Pàmpols per a dolmades | fulles de raïm | àcid | Europa SE | BL |
| Gundruk | fulles de raves, col i altres | àcid | Nepal | BL |
| Sinki | arrel de rave | àcid | Nepal | BL |
| Garri | arrel de mandioca | àcid | Àfrica W | BL, floridures, llevats |
| Sapal | tubercle de taro | àcid | Papua Nova Guinea | BL, llevats |
| Poi | tubercle de taro | àcid | Hawaii | BL |
| Tocosh | patata | àcid | Amèrica S | BL |
| Fufu | arrels de nyam, mandioca | àcid | Àfrica W | BL |
| Natto, Kinema i altres | faves de soia | àlcali | Japó, Àsia E | Bacillus subtilis (més info al meu post) |
| Salsa Gochujang | pebre, arròs, soja, cereals | àcid + dolç picant, condiment | Corea | Bacillus, Enterococcus, cianobacteri Aerosakkonema, floridures |
| Tempeh | faves de soia | àlcali | Indonèsia | Rhizopus |
| Salsa de soia | faves de soia | àlcali + àcid, condiment | Àsia E | Aspergillus oryzae (koji), BL, llevats |
| Miso | faves de soia, cereals | àlcali + àcid, condiment | Japó | Aspergillus oryzae (koji), BL, llevats |
| Oncom | restes de soia, mandioca i altres | àlcali | Indonèsia | Rhizopus, Neurospora |
| Sumbala, Dawadawa | grans de fabàcia néré | àlcali | Àfrica W | Bacillus, BL |
| Café (més info al meu post) | grans de cafè | àcid, beguda | Àfrica E | Enterobacteris, Bacillus, BL i llevats |
| Cacau | grans de cacau | àcid | Amèrica central i S | Llevats, BL i bacteris acètics |
| Olives de taula | fruits | àcid | Mediterrani | BL, llevats |
| Altres vegetals confitats | cogombres, albergínies, raves i altres | àcid | Mediterrani | BL, bacteris acètics |
| Pulque | saba de tija floral del maguei, Agave | alcohol, beguda | Mèxic | Zymomonas, BL, llevats |
| Vi | fruits de raïm | alcohol, beguda | Mediterrani | Llevats, i BL a malolàctica |
| Sidra | poma | alcohol, beguda | Europa W | Llevats |
| Sidra de pera | pera | alcohol, beguda | Regne Unit, França | Llevats |
| Vins de fruita | fruites diverses: cirera, banana i altres | alcohol, beguda | Europa N, Amèrica central | Llevats |
| Vinagre | fruites o cereals | àcid, condiment | Mediterrani | Bacteris acètics |
| Cerveses | grans de cereals | alcohol, beguda | Europa, Àsia W | Llevats |
| Cerveses “àcides”(més info al meu post) | grans de cereals | alcohol + àcid, beguda | Bèlgica, Alemanya | Llevats, BL |
| Kvass | grans de cereals | alcohol + àcid, beguda | Europa E | Llevats, BL |
| Sake, vi d’arròs | grans d’arròs | alcohol, beguda | Japó | Llevats, Aspergillus oryzae (koji) |
| Makgeolli, vi d’arròs coreà | grans de cereals | alcohol, beguda | Corea | Llevats, Aspergillus, BL, proteobacteris |
| Chicha | grans de moresc | alcohol, beguda | Amèrica S | BL, altres bacteris, llevats |
| Massa mare, sourdough | grans de cereals | àcid, massa | Europa, Àsia W, Amèrica N | BL, llevats |
| Appam | arròs, llet coco | àcid, massa | Índia | BL, llevats |
| Idli | arròs i llenties | àcid | Índia | BL |
| Kenkey | grans de moresc | àcid, massa | Àfrica W | BL, llevats |
| Pozol | grans de moresc, cacau | àcid, beguda | Amèrica central | BL, altres bacteris, llevats, floridures |
| Injera | grans de cereals (Eragrostis tef) | àcid, pa | Etiòpia, Àfrica E | BL, Bacillus, enterobacteris, llevats |
| Hidromel, Tej etíop | mel | alcohol, beguda | Àfrica, Àsia, Europa | Llevats |
| Formatges | llet | àcid | Mundial | BL, altres bacteris, llevats, floridures |
| Iogurt i semblants | llet | àcid | Europa E, Àsia W | BL |
| Crème fraiche | llet | àcid | França, Europa | BL |
| Quefir | llet | àcid | Caucas | BL, llevats |
| Kumis | llet d’euga | àcid, alcohol | Àsia central, Amèrica S | BL, llevats |
| Chal | llet de camella | àcid | Àsia central | BL, llevats |
| Leben | llet | àcid | Àfrica N, Àsia W | BL |
| Buttermilk | sèrum de mantega | àcid | Europa, Àsia W | BL |
| Embotits | carn de porc i altres | àcid | Europa | BL, llevats, floridures |
| Pernil | carn de porc | àcid | Europa | BL, altres bacteris, floridures |
| Nem chua | carn de porc, arròs, fulles plàtan | agredolç | Vietnam | BL |
| Satchu | carn | àcid | Himàlaia | BL, altres bacteris, llevats, floridures |
| Pemmican | carn de bisó, cèrvol i altres | àcid | Amèrica N | Diversos bacteris |
| Dodery | ossos animals | àcid | Sudan | Bacillus, altres bacteris, BL, llevats |
| Tiroi | musclos, altre marisc | àcid | Nova Zelanda | Diversos bacteris, BL |
| Kina | eriçons de mar | àlcali | Nova Zelanda | Diversos bacteris |
| Hákarl | carn de tauró | àlcali | Islàndia | Proteobacteris: Moraxella, Acinetobacter |
| Ngari | peix ciprínid | àcid | Índia, Himàlaia | BL, Bacillus, llevats |
| Surströmming | areng | àcid | Suècia, Europa N | Halanaerobium (arqueu), BL, altres bacteris |
| Nam-pla, bagoong i altres | peixos diversos | àcid, condiment | Àsia SE, Filipines, Europa | Bacillus, altres bacteris, arqueus halòfils |
| Garum | vísceres de peix | àcid, condiment | antigues Grècia, Roma, Bizanci | Diversos bacteris i arqueus |
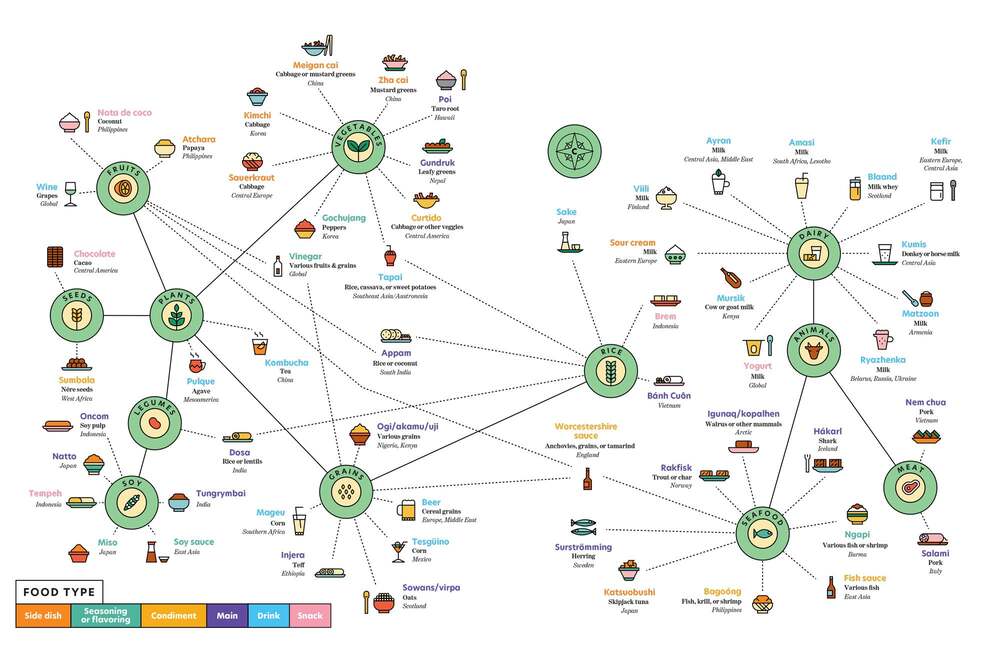
Els aliments fermentats són actualment una part important en la dieta humana a tot arreu, tant en regions on la seguretat alimentària i conservació actualment no estan ben controlades com en les més desenvolupades. És una tecnologia global entre els humans, i per tant és una prova de què prové dels primers humans. Com veiem (Taula 3), els substrats dels aliments poden ser vegetals, de les diferents parts de les plantes, o també de molt diversos animals.
A més, encara que les pràctiques culturals de fermentar aliments són molt variades globalment, sembla clar que en general els aliments fermentats ens agraden als humans. Aquesta preferència hauria emergit en paral·lel a una atracció adaptativa pels aromes i textures pròpies dels aliments fermentats per part dels primers humans. Per això podem observar com hi ha molts d’aquests aliments que són condiments, o sigui que s’afegeixen a altres aliments per tal de millorar la seva palatabilitat (Bryant et al 2023).
Aquesta gran diversitat dels aliments fermentats fa que alguns gustos i aromes ben estranys siguin molt apreciats per unes cultures i detestats per altres, com passa amb alguns formatges molt pudents, amb compostos volàtils amoniacals i de sofre. Hi ha una especificitat cultural en el seu consum. Els mateixos aromes que poden ser senyal de menjar “bo” en una cultura poden ser senyal de menjar dolent o passat en una altra. La capacitat per “degustar” menjars àcids, agres o amargs, gustos no habituals als aliments naturals i absents en altres animals, segurament ha evolucionat als humans amb la producció d’aliments fermentats (Frank et al 2022).
Com vèiem abans (Taules 1 i 2), el desenvolupament de la fermentació externa d’aliments va anar lligada a una pèrdua important de massa del còlon i de l’energia que s’hi produeix, i per tant això implica una reducció en la quantitat i diversitat de la microbiota intestinal perquè aquests no són tan necessaris. Això s’evidencia en fer anàlisis comparatives de la microbiota humana amb la dels altres homínids com ximpanzés, bonobos o goril·les (Moeller et al 2014).
D’altra banda, la preferència dels humans per als aliments fermentats també es demostra amb les anàlisis genètiques. Per exemple, alguns gens de receptors olfactius relacionats amb productes fermentats estan seleccionats positivament als humans i no als ximpanzés, com són els del metil-octanoat, d’olor afruitat produït pels llevats de vinificació, o del metilvalèric, aroma clau dels formatges madurats.
La capacitat de metabolitzar l’etanol produït a la fermentació alcohòlica i per tant poder consumir-lo en moderació és deguda a unes variants gèniques que codifiquen per a l’alcohol deshidrogenasa (ADH7), que lògicament s’haurien impulsat en els primers humans amb les primeres begudes obtingudes per fermentació. Tanmateix, sembla que aquesta capacitat seria prèvia als humans, ja que els altres grans primats la tenen, i fins i tot altres mamífers com alguns quiròpters. Tots ells són consumidors de fruites, que poden ser fermentades espontàniament a la natura, i per tant tots aquests animals haurien adquirit aquesta capacitat en consumir fruites que han estat parcialment fermentades (Janiak et al 2020). Així doncs els homínids ja estarien adaptats a metabolitzar l’etanol molt abans que els primers humans ho fessin de forma més dirigida (Carrigan et al 2015).
CONCLUSIÓ
Crec que aquesta hipòtesi de la fermentació externa dels aliments com a element clau en l’expansió del cervell observada en l’evolució des dels australopitecs als humans és molt versemblant. La fermentació dels aliments en molts casos és quasi espontània, inicialment requereix molt poca tecnologia i coneixements, i amb una mínima selecció dels productes resultants després de la fermentació, s’obtenen aliments més digeribles, i que es conserven millor, i que tenen gustos o textures nous i interessants.
Aquest desenvolupament dels aliments fermentats va permetre que no calgués tenir un volum considerable de còlon amb la seva microbiota tan diversa, per adquirir nutrients que es poden consumir elaborant-los prèviament. En reduir-se les necessitats calòriques del còlon, l’energia “sobrant” va poder ser dedicada cada cop més al cervell, facilitant-ne la seva expansió. Lògicament i en paral·lel, o en alguns casos posteriorment, els altres factors comentats com el consum de carn, noves tecnologies de cacera, socialització, i el foc, van permetre encara més aquest engrandiment del cervell, fins arribar als Homo sapiens.
Finalment, només vull comentar que aquest treball m‘ha agradat especialment perquè tot plegat lliga tres dels temes que més m’atrauen científicament:
1) Aliments fermentats, o tal com a l’article de Bryant et al es diu, “fermentació externa”. De fet aquest denominació d’externa m’ha sorprès, ja que no havia pensat mai en anomenar “fermentació interna” al conjunt de processos de modificació o degradació o síntesi de compostos que du a terme la microbiota intestinal. Però vaja, és veritat. En qualsevol cas, els aliments fermentats i els aspectes dels beneficis dels microorganismes (“els microbis bons”) han estat sempre el meu tema primordial de treball de recerca, i de docència, i d’interès des que vaig acabar la llicenciatura de Biologia, ara fa 50 anys.
2) Microbiota intestinal. Des de fa uns quants anys és un tema que m’interessa molt. Encara que no l’he treballat directament a nivell de recerca, sí que he anat coneixent-lo, i el toco una mica a nivell docent. Com estem veient aquests darrers anys el paper de la microbiota intestinal en el manteniment saludable del cos és molt més important del que ens pensàvem, si bé encara falta molt per conèixer. Curiosament i una mica decebut, he descobert amb aquest treball, que els humans hem prescindit una mica de la microbiota intestinal i l’hem reduït en comparació als altres primats, justament amb el desenvolupament d’aquesta “fermentació externa”.
3) Origen i evolució humana. Per descomptat, aquest tema m’interessa molt, com suposo que a tothom. Amb un cert coneixement dels sers vius i admirant com funciona tota l’evolució biològica, saber més de com va aparèixer la nostra espècie i les properes, és apassionant.
BIBLIOGRAFIA
Alexander C, Swanson KS, Fahey GC, Garleb KA (2019) Perspective: physiologic importance of short-chain fatty acids from nondigestible carbohydrate fermentation. Adv Nutr 10, 576–589
Amato KR, Chaves OM, Mallott EK et al (2021) Fermented food consumption in wild nonhuman primates and its ecological drivers. Am J Phys Anthropol 175, 513–530
Bryant KL, Hansen C, Hecht EE (2023) Fermentation technology as a driver of human brain expansion. Commun Biol 6, 1190
Bunn HT, Ezzo J (1993) Hunting and Scavenging by Plio-Pleistocene Hominids: Nutritional Constraints, Archaeological Patterns, and Behavioural Implications. J Archaeol Sci 20, 365-398
Carrigan MA, Uryasev O, Frye CB et al (2015) Hominids adapted to metabolize ethanol long before human-directed fermentation. Proc Natl Acad Sci USA 112, 458–463
Cordain L, Eaton S, Miller J et al. (2002) The paradoxical nature of hunter-gatherer diets: meat-based, yet non-atherogenic. Eur J Clin Nutr 56, S42–S52
Dyer R (2023) Food preserving technique may have sparked human brain growth, scientists say. Science Alert – Humans, 3/12/2023
Frank HER, Amato K, Trautwein M et al. (2022) The evolution of sour taste. Proc. Biol. Sci. 289, 20211918
Goren-Inbar N, Alperson N, Kislev ME et al. (2004) Evidence of Hominin Control of Fire at Gesher Benot Ya`aqov, Israel. Science 304,725-727
Hlubik S, Cutts R, Braun DR et al (2019) Hominin fire use in the Okote member at Koobi Fora, Kenya: New evidence for the old debate. J Human Evol 133, 214-229
Janiak MC, Pinto SL, Duytschaever G et al (2020) Genetic evidence of widespread variation in ethanol metabolism among mammals: revisiting the ‘myth’ of natural intoxication. Biol. Lett. 16, 20200070
Mann N (2000) Dietary lean red meat and human evolution. Eur J Nutr 39, 71–79 (2000)
Marean C, Bar-Matthews M, Bernatchez J. et al (2007) Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature 449, 905–908
McClave SA, Snider HL (2001) Dissecting the energy needs of the body. Curr Opin Clin Nutr Metab Care 4(2):143-7
Miller IF, Barton RF, Nunn CL (2019) Quantitative uniqueness of human brain evolution revealed through phylogenetic comparative analysis. eLife 8:e41250
Moeller AH, Li Y, Ngole EM et al (2014) Rapid changes in the gut microbiome during human evolution. Proc Natl Acad Sci USA 111, 16431–16435
O’Hara AM, Shanahan F (2006) The gut flora as a forgotten organ. EMBO Rep 7, 688–693
Padmaja G, Steinkraus KH (1995) Cyanide detoxification in cassava for food and feed uses. Crit Rev Food Sci Nutr 35, 299–339
Ponce de León MS, Bienvenu T, Marom A et al. (2021) The primitive brain of early Homo. Science 372, 165-171
Ohland CL, Macnaughton WK (2010) Probiotic bacteria and intestinal epithelial barrier function. Am J Physiol Gastrointest Liver Physiol 298, G807–19
Sandhu KS, Punia S, Kaur M (2017) Fermentation of cereals: a tool to enhance bioactive compounds. Plant biotechnology: Recent advancements and developments 157, 157–170
Speth JD (2019) Neanderthals, vitamin C, and scurvy. Quat. Int. 500, 172–184
Wrangham RW, Jones JH, Laden G et al. (1999) The Raw and the Stolen: Cooking and the Ecology of Human Origins. Curr Anthrop 40:5, 567-594
