Arxiu del Blog

Trichodesmium, el fixador de nitrogen més important al mar, i molt més
14 juny 2023
Click here for the English version: Trichodesmium is the most important nitrogen-fixing cyanobacteria
El gènere Trichodesmium és un dels cianobacteris més importants des de diversos punts de vista: és prolífic als oceans, hi fixa la meitat del nitrogen atmosfèric necessari per a tota la cadena alimentària, és fotosintètic, és colonial i forma flocs que es desplacen verticalment entre la superfície i fins a 200 m, contribuint als cicles biològics del nitrogen, carboni, fòsfor i del ferro.
ELS CIANOBACTERIS SÓN ORGANISMES CLAU A L’EVOLUCIÓ I A L’ECOLOGIA DE LA TERRA
Abans eren coneguts com a “algues verd-blaves” o cianofícies, per la seva semblança amb les algues, essent fotosintètics autòtrofs i d’aspectes filamentosos, i encara se’ls anomena vulgarment com un tipus de microalgues. Tanmateix, els cianobacteris són procariotes, i per tant no tenen res a veure estructuralment i filogenèticament amb les algues, ja que aquestes són eucariotes.
Són un dels principals fílums dels bacteris, dins el supertàxon Terrabacteria. Vegeu al respecte el meu article sobre els principals fílums bacterians. De fet, són uns dels bacteris més antics, amb fòssils de fa 3500 milions d’anys. Els cianobacteris van “inventar” la fotosíntesi oxigènica, per fotofosforilació no cíclica, amb 2 fotosistemes i clorofil·la, a diferència d’altres bacteris fotosintètics anoxigènics, amb només un fotosistema i bacterioclorofil·les. L’oxigen que anaren produint els cianobacteris, sobretot entre fa 2400 i 2000 milions d’anys, va generar l’atmosfera actual amb un 21% O2. I de fet ells són l’origen evolutiu dels primers cloroplasts i altres plastidis presents a algues i plantes, els eucariotes que aparegueren després, ja que els cianobacteris foren incorporats per endosimbiosi als primers eucariotes.
Són gramnegatius i tenen membranes internes, anomenades tilacoides, on s’acumulen els pigments i on té lloc la fotosíntesi. Són un fílum gran i molt divers, amb molts tipus de formes, tant unicel·lulars, como colonials i moltes filamentoses (Figura 1). Els cianobacteris prosperen en molt diversos hàbitats per tota la Terra i són els principals contribuïdors als cicles biogeoquímics globals. Molts d’ells a més de ser fotosintètics, són fixadors de N2 atmosfèric —diazòtrofs—, convertint-lo a amoni i/o nitrits i nitrats. Alguns fan la fixació de nitrogen en cèl·lules especialitzades, com els heterocists d’Anabena i altres gèneres (Figura 1). Hi ha molts fixadors de N2 de vida lliure però també n’hi ha de simbiòtics amb les arrels de les plantes, anomenats cianobionts, com el mateix Anabaena.
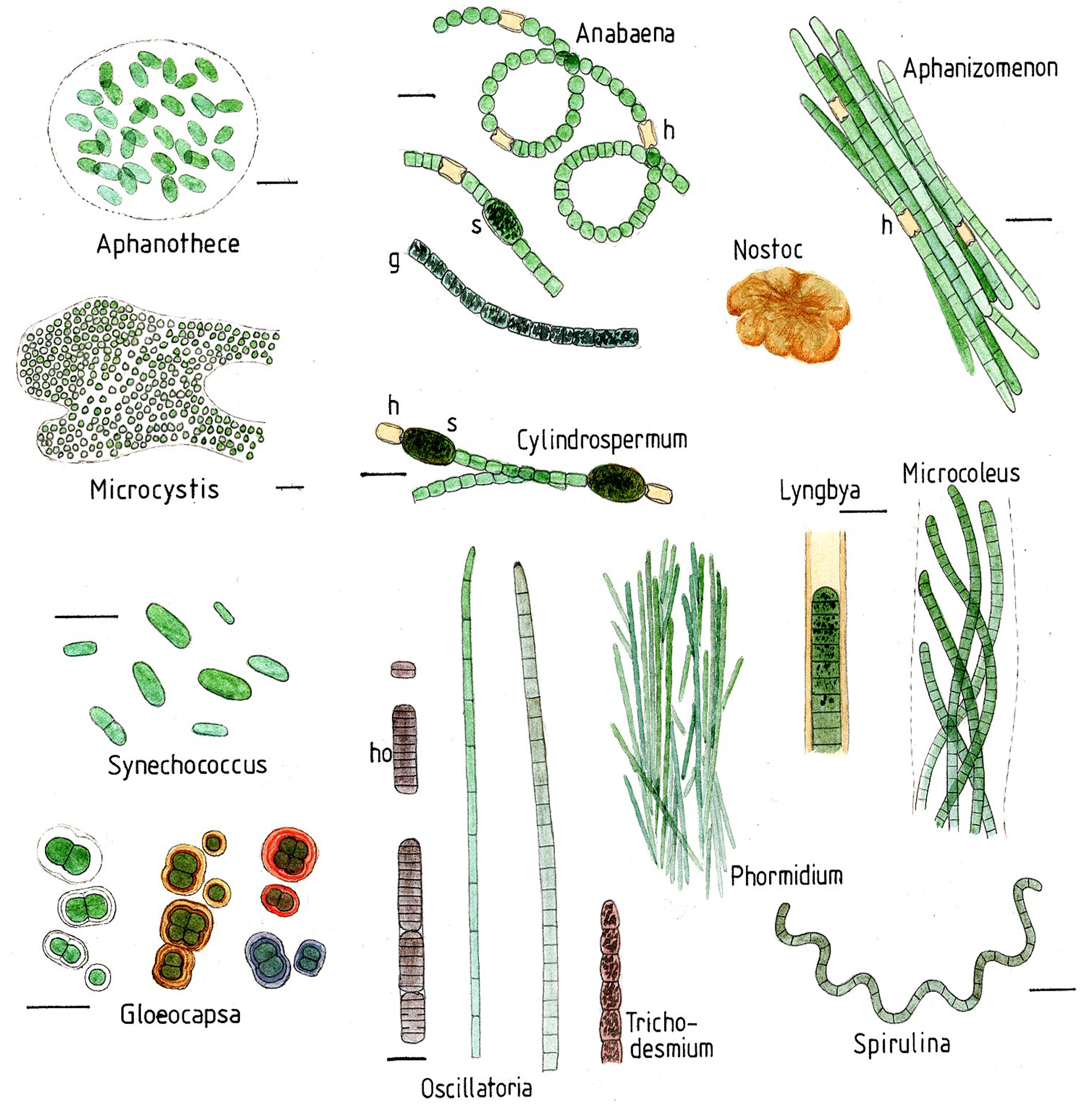
A banda de trobar-se en ambients terrestres i aigües superficials, els cianobacteris són ubics als ambients marins, on tenen un paper important com a productors primaris, part del fitoplàncton. Els principals cianobacteris de mar obert són Crocosphaera, Trichodesmium, Synechococcus i Prochlorococcus. Aquests 2 darrers gèneres són dels anomenats picobacteris, perquè són molt petits, de només 0.5-0.8 μm, i malgrat la seva mida són els organismes més abundants a la Terra: uns 105 bacteris per cada mL d’aigua marina, sobretot a totes les regions oligotròfiques (amb pocs nutrients) dels oceans. Es calcula que només els Prochlorococcus produeixen el 20% de l’oxigen atmosfèric (Partensky et al 1999).
Els cianobacteris marins també són presents a l’anomenat aeroplàncton atmosfèric que es forma amb les esquitxades de les onades, mitjançant el qual aquests bacteris són transportats a altres regions. Alguns cianobacteris poden formar proliferacions algals nocives (blooms) —per excés de nutrients de fertilitzants o residus amb altes temperatures— que al seu torn provoquen desequilibris de l’ecosistema i poden produir toxines, les cianotoxines (Huisman et al 2018).
Biotecnològicament, els cianobacteris són importants com a organismes model d’estudi dels mecanismes fotosintètics i potencials aplicacions. A més, són coneguts —a banda d’alguns productes del metabolisme secundari— per la seva utilització com a menjar, o sigui l’anomenada “proteïna unicel·lular” (Single Cell Protein, SCP) o proteïna microbiana. Gèneres com Arthrospira (abans Spirulina) o Aphanizomenon són utilitzats com a suplements dietètics i tenen efectes positius sobre la salut. Ho podeu veure al meu article sobre la carn alternativa.
TRICHODESMIUM
És un gènere de cianobacteris filamentosos, que es troba en aigües oceàniques tropicals i subtropicals pobres en nutrients, com al mar Roig i als mars d’Austràlia, on fou descrit en primer lloc al segle 18 per l’explorador anglès capità James Cook. De fet, el mar Roig té aquest nom pel color rogenc que s’observa a vegades degut a la proliferació de T. erythraeum i altres espècies semblants. T. erythraeum és la més estudiada i la única seqüenciada fins ara d’aquest gènere. El seu genoma de 7.75 Mb és un dels més grans de tots els bacteris seqüenciats. Una altra espècie ben coneguda per les proliferacions en superfície és T. thiebautii.
Els filaments de Trichodesmium són cadenes d’unitats cel·lulars separades per septes (Figura 2). Els filaments s’anomenen tricomes —mateix nom (pèls en grec) que els fins apèndixs o pèls de plantes i algues. Els tricomes s’agrupen formant flocs de fins a 2 mm, visibles a simple vista (Figura 3), i la seva acumulació a la superfície marina en forma de bandes és visible des de l’espai (Figura 4). Per aquest aspecte també són anomenats “serradures marines” o també “camins de sorra”. De fet, el nom d’aquest gènere prové del grec Trichos = pèls, i Desmos = bandes, o sigui bandes de pèls.
Les colònies de Trichodesmium són un substrat preferent de nombrosos altres organismes oceànics, incloent altres bacteris, protistes com diatomees, dinoflagel·lats, protozous, i sobretot els copèpodes (crustacis, animals). Per tant, aquest gènere és el pilar substancial de complexos microambients marins.



———————
LA FIXACIÓ DE NITROGEN PER TRICHODESMIUM
La recerca feta en diversos estudis els darrers anys ha fet palesa la substancial contribució del cianobacteri diazòtrof Trichodesmium a la captació de nitrogen atmosfèric als ecosistemes marins globals i la cabdal importància ecològica d’aquest gènere (Capone et al 1997; Bergman et al 2013).
Amb la seva gran capacitat de fixar N2 atmosfèric, Trichodesmium és el principal diazòtrof entre els cianobacteris i també el principal organisme diazòtrof dels sistemes pelàgics marins. És una font important de “nou” nitrogen a les aigües pobres en nutrients que habita, produint la meitat del nitrogen que cal per a la producció primària als oceans (MicrobeWiki 2010). L’aportació global de fixació de nitrogen per Trichodesmium és d’aproximadament 60-80 Tg —Teragrams (1012 g), o sigui megatones— de N per any.
Com sabeu, la fixació biològica de nitrogen atmosfèric només la poden fer bacteris, i consisteix en la reducció de N2 —molt inert, cal molta energia per reduir-lo— a 2 molècules de amoni mitjançant un complex enzimàtic, la dinitrogenasa, en un seguit de reaccions semblants a una cadena de transport d’electrons amb piruvat com a donador d’e–, on intervenen una flavodoxina oxidoreductasa i el complex dinitrogenasa pròpiament, amb requeriment d’uns 24 mols ATP per mol N2 fixat. Com que la nitrogenasa és inhibida per l’oxigen, els organismes diazòtrofs tenen diversos mecanismes de poder fixar el nitrogen sense la presència d’oxigen:
- Ser anaeròbics, com alguns clostridials i també alguns fotosintètics anoxigènics.
- Aeròbics que consumeixen ràpidament l’O2 —anomenada protecció respiratòria—, com Azotobacter, que té una citocrom-oxidasa molt activa, i així no hi ha oxigen lliure.
- Aeròbics amb cèl·lules especialitzades que contenen la nitrogenasa, com els heterocists de molts cianobacteris (ex. Anabaena, Figura 1) que impedeixen l’entrada d’oxigen. A més, donat que els cianobacteris són fotosintètics oxigènics, estan produint oxigen, i per tant, cal tenir la nitrogenasa aïllada en aquests heterocists, que fixen nitrogen i no fan fotosíntesi.
- Aeròbics simbiòtics de plantes, com Rhizobium dels nòduls de lleguminoses on el compost leghemoglobina segresta l’oxigen.
Es dona la paradoxa que Trichodesmium, malgrat ser un cianobacteri fotosintètic oxigènic, i per tant produir oxigen i ser fixador de N2, no té heterocists, si bé no és l’únic gènere amb aquestes característiques (Bergman et al 1997). Enlloc dels heterocists, la fixació de nitrogen a Trichodesmium té lloc a unes cèl·lules relativament especialitzades, els diazòcits, que contenen la nitrogenasa però no tenen parets cel·lulars engruixides (Figura 5). Com veiem, la zona de diazòcits no té grànuls, és més clara, es tenyeix diferent de la resta i on es detecta per immunofluorescència la presència de la proteïna NifH de la nitrogenasa (Bergman et al 2013).

En els cianobacteris amb heterocists, la fixació de nitrogen i la fotosíntesi es produeixen simultàniament, durant el dia amb llum, perquè els dos processos se separen en l’espai utilitzant els heterocists per protegir l’enzim nitrogenasa sensible de l’oxigen. L’energia que cal als heterocists és aportada per les altres cèl·lules que realitzen la fotosíntesi. Alguns altres cianobacteris, els menys, fan la separació en el temps fent fotosíntesi durant el dia i fixant N2 per la nit.
En canvi, les zones de diazòcits de Trichodesmium no tenen la protecció estructural dels heterocists i la quasi inexistent separació temporal entre els dos processos és peculiar i única entre els diazòtrofs. Així, la nitrogenasa dels diazòcits és activa durant les hores de llum, però sobretot amb un màxim al migdia, coincidint amb una baixada en la captació de CO2 i la producció d’O2, i un augment dels mecanismes segrestadors d’O2. Aquests serien d’una banda l’esmentada protecció respiratòria —amb augment de l’activitat citocrom c oxidasa—, i a més l’anomenada reacció de Mehler, també observada als cloroplasts de plantes, que és la reducció de l’O2 a H2O2 per electrons del fotosistema I, on aquest és acoblat reversiblement al fotosistema II. Tot seguit el H2O2 és reduït per algun dels sistemes antioxidants propis. Per tant, sorprenentment la fixació de N2 depèn de l’activitat dels fotosistemes a Trichodesmium (Bergman et al 2013).
———————
ELS AGREGATS DE TRICHODESMIUM
Aquest cianobacteri es troba als oceans o bé en forma de filaments multicel·lulars dispersos, o bé en forma d’agregats que comprenen centenars de filaments, amb l’aspecte final comentat de flocs o colònies (Figures 3 i 5a). L’agregació de filaments passa sobretot durant el dia amb llum, després de la fase exponencial de creixement, i també en situacions d’estrès per canvis en les condicions ambientals, com la mancança de fosfat o ferro. Els agregats, com a flocs que són, permeten una ràpida migració vertical cap al fons, sobretot per tal de capturar fosfat. A la superfície assimilen i acumulen C i N i un cop al fons, uns 100-200 m, capturen el fòsfor que manca a la superfície (Rodríguez 2017). Els agregats alhora creen microambients subòxics que redueixen la fixació de N2 i de CO2, però també faciliten la captació de ferro que alhora permet recuperar la fixació de N2 simultània amb la fotosíntesi (Pfreundt et al 2023).
Recentment s’ha vist que la formació dels agregats a partir dels filaments individuals pot ser molt ràpida i deguda a canvis bruscos en la llum com pot ser el pas de núvols (Figura 6), i que les agregacions són degudes a canvis en la motilitat dels filaments individuals, per tal de disposar-se en contacte i solapant-se uns amb els altres, el que es coneix com a tigmotàxia o orientació per contacte (Figura 5a). Quan dos filaments es toquen, rellisca un damunt de l’altre, i en direcció contrària un de l’altre per tal d’anar augmentant la mida de l’agregat, i quant més freqüents són les reversions, més dens és el floc (Pfreundt et al 2023).

———————
NO NOMÉS EL NITROGEN, TAMBÉ EL FERRO I EL CARBONI
La biodisponibilitat del ferro (Fe) limita el creixement del fitoplàncton a mar obert dels vastos oceans. La pols transportada per l’atmosfera des dels deserts a la superfície dels oceans és rica en Fe però aquest és poc biodisponible perquè és poc soluble en aigua de mar i la pols s’enfonsa ràpidament per sota de la zona fòtica on és el fitoplàncton.
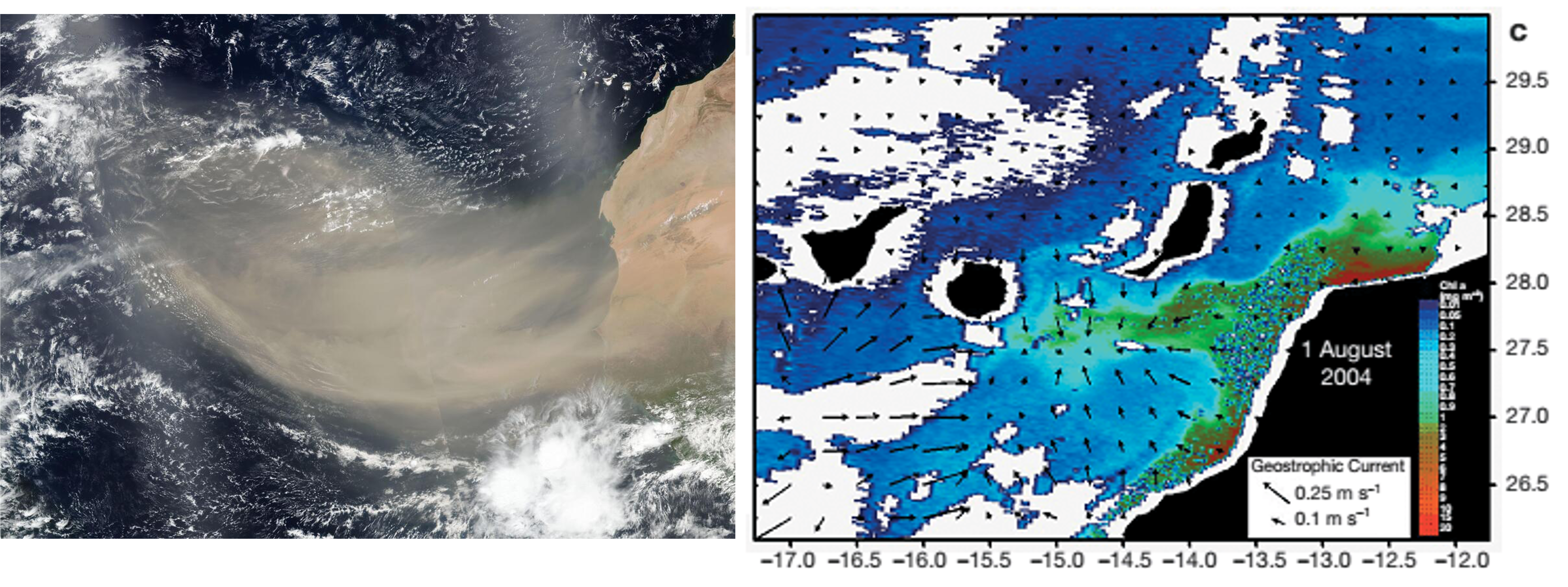
L’excepció es Trichodesmium, perquè les seves colònies o flocs capturen les partícules d’aquesta pols i simbiòticament amb altres bacteris associats promouen la dissolució de la pols i la captació del Fe mitjançant sideròfors (Figura 7) (Basu et al 2019). En quant a la contribució de la pols del Sàhara a l’oceà Atlàntic s’ha demostrat la relació amb la proliferació de Trichodesmium al voltant de les Canàries (Figura 8) (Ramos et al 2005).
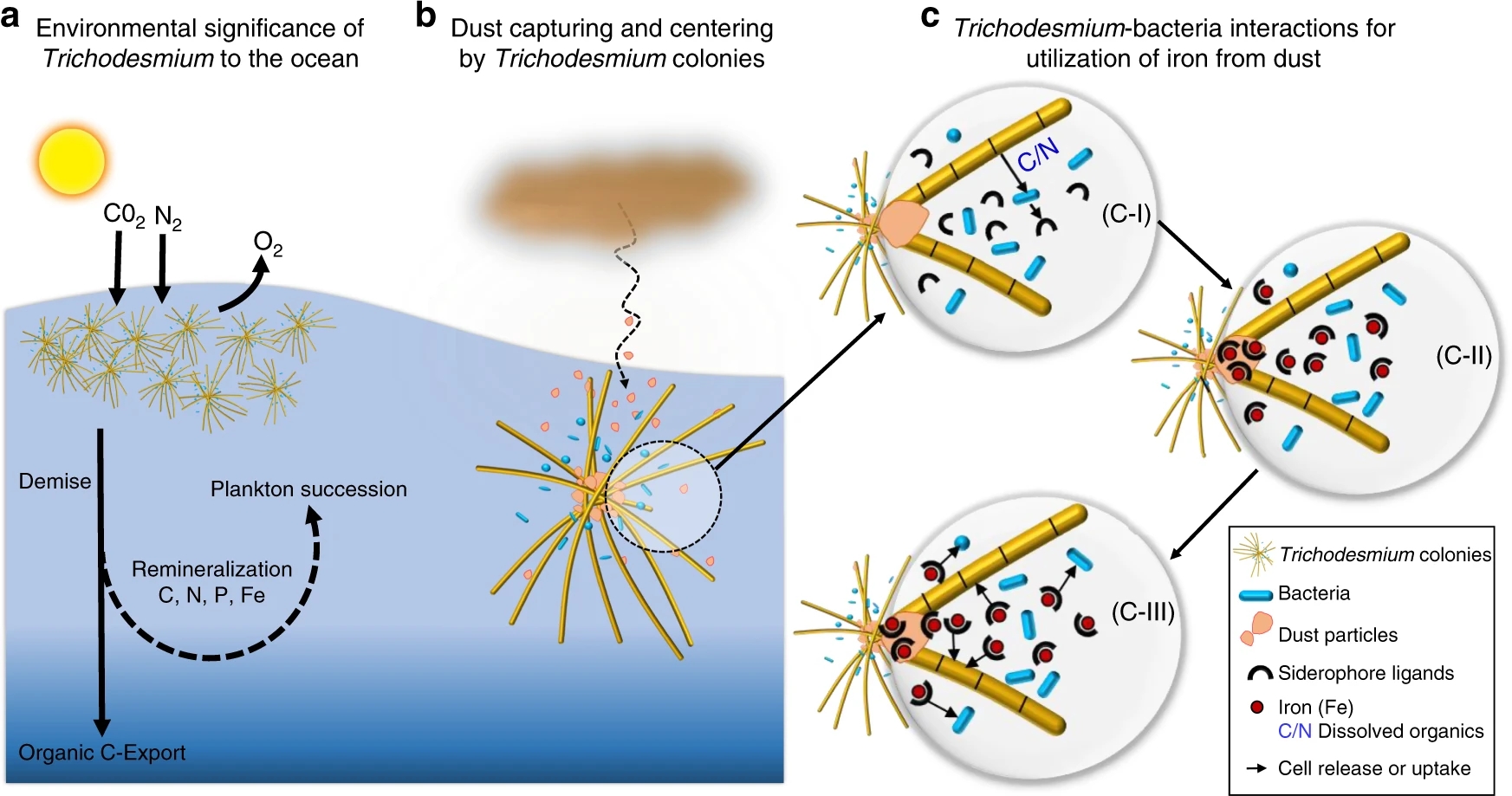
I a més a més, el cianobacteri Trichodesmium contribueix directament en l’exportació i segrest de carboni al llit marí. Fins ara se sabia que aquests microorganismes reciclaven el CO₂ captat de l’atmosfera mitjançant la fotosíntesi, però no que també se l’enduien amb ells a les profunditats quan moren i s’enfonsen, com sí que ho fa la resta del fitoplàncton, és a dir, els eucariotes. Això s’anomena la bomba biològica de carboni, i d’aquesta manera s’emmagatzema al llit marí aproximadament el doble de carboni del que es troba actualment a l’atmosfera. Fins ara això s’atribuïa quasi exclusivament al fitoplàncton eucariota, i els models biogeoquímics globals, és a dir, les eines que s’utilitzen per fer prediccions sobre l’evolució i els fluxos de carboni al planeta, no tenen en compte la contribució directa dels diazòtrofs com Trichodesmium en aquest procés (Cornejo 2022, Bonnet 2022).
———————
CONCLUSIÓ
Per concloure, es pot dir que el cianobacteri Trichodesmium és un dels organismes més abundants dels ambients marins, és part substancial del fitoplàncton i per tant molt important productor primari. És el fixador de nitrogen més rellevant al mar, i un dels més importants per als cicles biogeoquímics globals de nitrogen, carboni, fòsfor i ferro. I a més, té una variable i complexa estructura d’agrupaments, i és encara poc conegut del punt de vista de funcionament del seu metabolisme i de regulació del seu gran genoma, malgrat ser procariota.
———————
BIBLIOGRAFIA
Basu S et al (2019) Colonies of marine cyanobacteria Trichodesmium interact with associated bacteria to acquire iron from dust. Comm Biol 2, 28
Bergman B et al (1997) N2 fixation by non-heterocystous cyanobacteria. FEMS Microbiol Rev 19, 139–185
Bergman B et al (2013) Trichodesmium –a widespread marine cyanobacterium with unusual nitrogen fixation properties. FEMS Microbiol Rev 37, 286–302
Bonnet S et al (2022) Diazotrophs are overlooked contributors to carbon and nitrogen export to the deep ocean. The ISME Journal, 1-12
Capone DG et al (1997) Trichodesmium, a Globally Significant Marine Cyanobacterium. Science 276, 1221-1229
Cornejo FM (2022) Els bacteris marins diazòtrofs, petits grans aliats contra el canvi climàtic. Institut de Ciències del Mar ICM-CSIC, 18 oct. 2022
Huisman et al (2018) Cyanobacterial blooms. Nature Rev Microbiol 16, 471-483
Le Page M (2023) Ocean-fertilising bacteria work together to adapt to light levels. New Scientist, 25 May 2023
MicrobeWiki (2010) Trichodesmium. A Microbial Biorealm page, Kenyon College, 6 Aug 2010
Partensky F et al (1999) Prochlorococcus, a marine photosynthetic prokaryote of global significance. Microbiol Mol Biol Rev 63, 106-127
Pfreundt U et al (2023) Controlled motility in the cyanobacterium Trichodesmium regulates aggregate architecture. Science 380,830-835
Ramos AG et al (2005) Bloom of the marine diazotrophic cyanobacterium Trichodesmium erythraeum in the Northwest African Upwelling. Marine Ecol Progress Series 301, 303-305
Rodríguez F (2017) Más respuestas sobre Trichodesmium. Blog Fitopasión, 1 agosto 2017
Wikipedia contributors (2023, May 28). Cyanobacteria. Wikipedia, The Free Encyclopedia
Wikipedia contributors (2022, July 18). Trichodesmium. Wikipedia, The Free Encyclopedia
WHOI, Woods Hole Oceanographic Institution (2023) The many faces of Trichodesmium

Microbiota de la hibernació: els bacteris treballen mentre l’animal dorm
13 abril 2023
Click here for the English version: Hibernation microbiota
HIBERNACIÓ
La hibernació és un estat de mínima activitat i alentiment del metabolisme que duen a terme alguns animals —bàsicament els mamífers—, usualment durant l’hivern, encara que n’hi ha alguns que fan “estivació”. Quan hibernen, els baixa la temperatura, les freqüències cardíaca i de respiració, i el metabolisme en general. Aquest estat corporal fisiològic es coneix com a torpor.
Per exemple, els esquirols de terra (Figura 1 ) o les marmotes hibernen uns 6 mesos, de la tardor a la primavera, respirant un cop per minut, amb uns 5 batecs cardíacs per minut i la temperatura els baixa a només 5-10oC. Aquest estat de torpor no és constant, sinó que cada 10-12 dies hi ha unes fases d’interrupció, on reviuen una mica durant unes 12 hores, els puja la temperatura (Figura 2), tremolen una mica com per escalfar-se i el metabolisme s’activa (Carey et al. 2003), però ni mengen ni beuen ni defequen (Wilke 2020).

Figura 1. Un esquirol de terra (Ictidomys tridecemlineatus) hibernant. Imatge de R. Streiffer, treta de Garcia de Jesús (2022).
———————
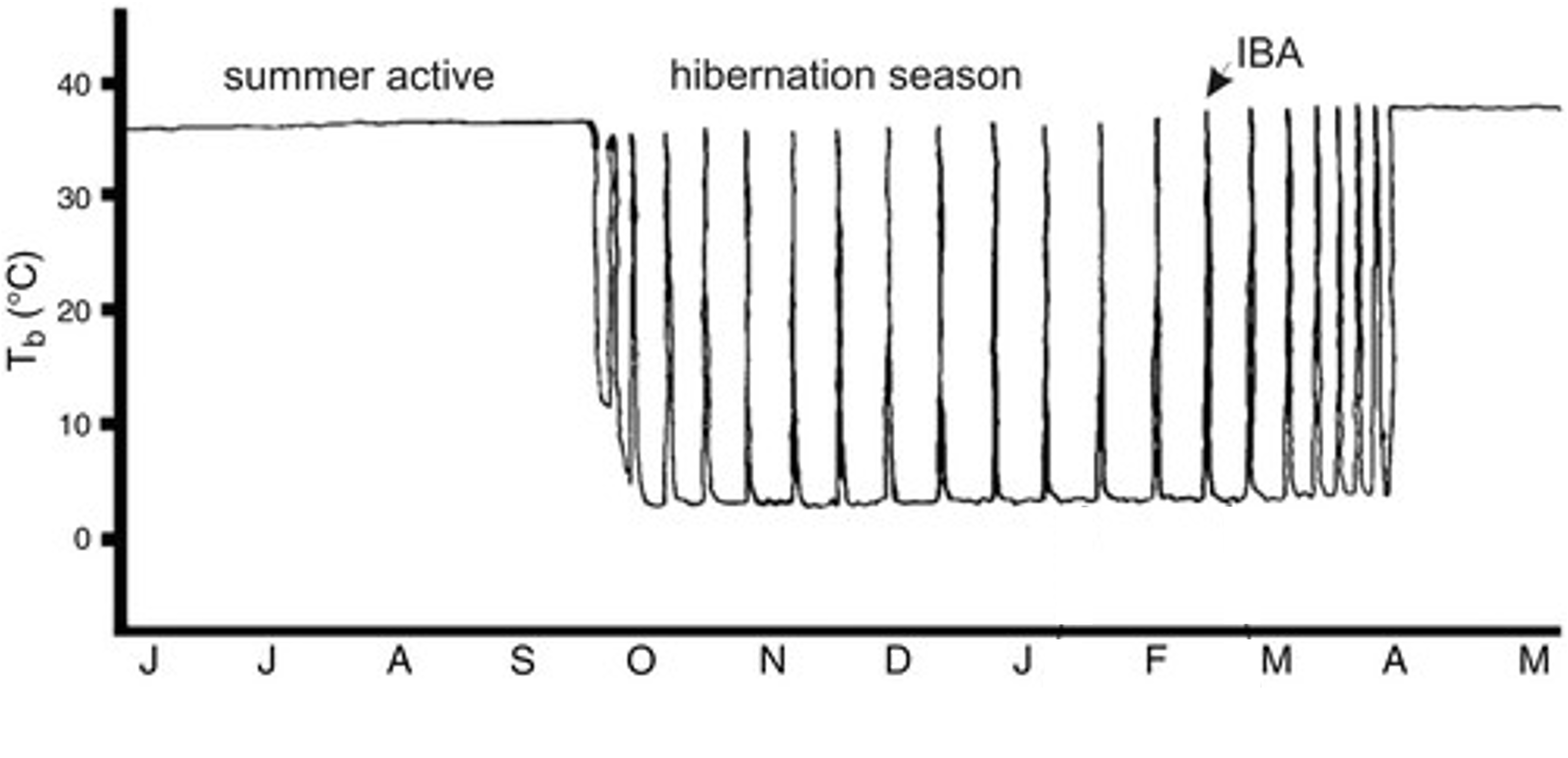
Figura 2. Temperatura corporal (Tb) d’un esquirol de terra al llarg de l’any, de juny a maig. Al període d’hibernació s’observen els cicles de llargues fases de torpor (10-12 dies) amb les petites fases d’interrupció (IBA, 12 hores) (Adaptat de Carey et al. 2003).
La hibernació serveix per mantenir i estalviar l’energia corporal quan no hi ha prou menjar disponible. Per això, abans d’hibernar i en general tots els mesos més càlids, aquests animals mengen molt, acumulant suficient reserves energètiques per al període hivernal. Tots ells hibernen en caus o coves, resguardats una mica del fred i altres inclemències exteriors.
El període d’hibernació és molt variable, des de poques setmanes a mesos, i pot ser obligat o facultatiu. També es distingeixen diferents nivells de hibernació, des d’hivernants profunds com molts rosegadors, el lèmur ratolí, l’eriçó i força marsupials, fins als hivernants més lleugers com els óssos. En aquests la temperatura corporal només els baixa uns 5oC i en canvi en alguns rosegadors baixa més de 30oC.
D’altra banda, els animals vertebrats ectotèrmics —de sang freda— com peixos, amfibis i rèptils, també baixen la seva activitat metabòlica i la temperatura corporal i entren en un estat de torpor, però no es considera hibernació perquè no ho fan de forma activa sinó que és en funció de l’ambient extern.
———————
METABOLISME DE LA HIBERNACIÓ
La hibernació és una estratègia dels mamífers que utilitza la plasticitat metabòlica per reduir les demandes d’energia i permetre dejunis de llarg termini. Per permetre aquests llargs períodes sense menjar, molts d’aquests animals mengen molt més abans d’hibernar, i acumulen greix corporal. Els óssos bruns per exemple arriben a doblar els dipòsits de greix i van consumint els lípids guardats al llarg de la hibernació. Encara que aquests óssos esdevenen obesos estacionalment, es mantenen metabòlicament sans, la qual cosa contrasta amb la forta relació entre obesitat i resistència a la insulina i altres problemes metabòlics que s’observen en els humans (Sommer et al. 2016).
El dejuni mitiga l’escassetat d’aliments hivernal però s’elimina el nitrogen de la dieta, amb la qual cosa posa en perill l’equilibri corporal de proteïna (Regan 2022). L’atròfia muscular deguda al dejú —o a la inactivitat— allibera compostos de nitrogen com l’amoni, que poden ser tòxics, i el cos n’ha d’eliminar l’excés, normalment en forma d’urea, mitjançant l’orina. Tanmateix, els animals hivernants com els molt estudiats esquirols de terra mantenen o perden molt poca massa muscular al llarg de l’hivern, i fins i tot a la fase final de la hibernació la taxa de síntesi de proteïnes augmenta fins a nivells semblants a quan estaran actius. Després veurem el paper de la microbiota en aquesta recuperació del nitrogen.
El coneixement de com aquests animals mantenen la massa muscular pot ajudar a trobar remei en el cas de les persones amb pèrdua muscular degut a malnutrició, sedentarisme forçat o malalties de desgast muscular (García de Jesús 2022).
———————
LA MICROBIOTA INTESTINAL DELS HIBERNANTS
Un dels mamífers hivernants més coneguts i estudiats és l’ós bru (Ursus arctos). Sommer et al. (2016) van fer un estudi amb 16 óssos bruns euroasiàtics en llibertat als boscos del centre i nord de Suècia, prenent-los mostres fecals i de sang per a l’estudi metabòlic, tant durant la hibernació (febrer-març) com al període actiu (juny). La microbiota fecal fou analitzada per metagenòmica, extraient el DNA genòmic, amplificant la regió V2-V3 del gen per al 16S rRNA, seqüenciant-la per 454 piroseqüenciació i analitzant-ho amb el programa MacQIIME per obtenir-ne la identificació. Els resultats observats van ser una clara diferència entre hivern i estiu, demostrable amb una anàlisi PCA (Figura 3, esquerra). També veieren una major diversitat de microbiota a l’estiu, 4 cops respecte la d’hivern, la qual era més homogènia que la d’estiu (resultats no mostrats). La heterogeneïtat de l’estiu pot estar relacionada amb una dieta més variada. La menor diversitat hivernal va lligada a una reducció sobretot de Firmicutes i d’Actinobacteria, i en canvi hi ha un augment de Bacteroidetes (Figura 3, dreta).
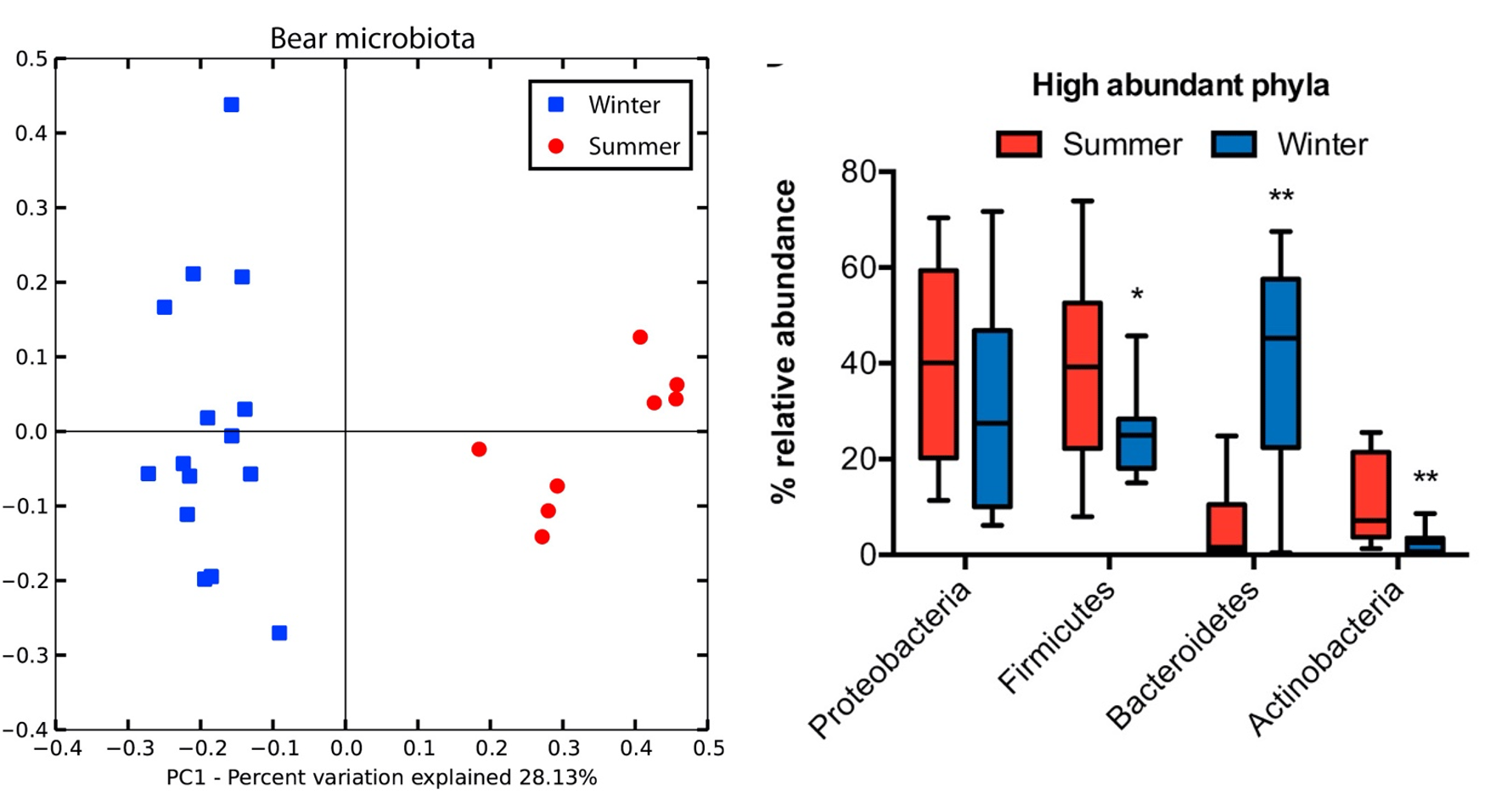
Figura 3. Microbiota fecal de l’ós bru comparada a l’hivern (blau) i l’estiu (vermell). Esquerra: PCA (anàlisi de components principals); dreta: abundància relativa dels fílums més abundats (Sommer et al. 2016).
L’augment de Bacteroidetes —sobretot Bacteroides fragilis— durant la hibernació pot explicar-se per llur capacitat de degradar glicans de l’hoste en absència de polisacàrids de la dieta i també perquè els Bacteroidetes poden metabolitzar proteïnes i greixos de l’epiteli intestinal. Els Firmicutes com Streptococcus probablement disminueixen perquè requereixen més fibra, la de la dieta (Sommer et al. 2016).
S’ha vist que la hibernació dels óssos bruns també afecta al metabolisme, sobretot lipídic. I a més, en fer transplantaments de la microbiota dels óssos hivernants o dels actius al juny a ratolins sense microbiota comporta que els ratolins adquireixen els trets metabòlics estacionals corresponents respectivament d’hivern o estiu (Sommer et al. 2016).
Cal senyalar que les temperatures corporals més baixes durant la hibernació són molt per sota les temperatures òptimes de la majoria de microbis intestinals, però com he comentat abans, el torpor és interromput periòdicament per unes curtes fases de pujada tèrmica que permeten —a banda d’activar el metabolisme de l’animal— incrementar el metabolisme bacterià, amb la conseqüent degradació dels substrats presents al lumen intestinal (Carey et al. 2013).
Uns dels animals més estudiats en relació a la hibernació i llur microbiota són els esquirols de terra, i en concret l’anomenat suslic (de l’original suslik en rus) o esquirol de terra de 13 bandes, Ictidomys tridecemlineatus (abans Spermophilus tridecemlineatus) (Figura 4). Tenen una coloració llampant, amb ratlles i taques al pelatge dorsal que fan que sigui molt fàcil distingir-los d’altres espècies. Aquests esquirols de terra, junt amb els més coneguts arborícoles, les marmotes, els esquirols voladors i els gossets de les praderies, constitueixen la família d’esciúrids, dins l’ordre Rodentia, els rosegadors.

Figura 4. EL suslic o esquirol de terra de 13 bandes (Ictidomys tridecemlineatus). Imatge de Cheryl Gorske, treta de Pinterest.
L’efecte del cicle anual d’hibernació en la microbiota intestinal del suslic ha estat estudiat (Carey et al. 2013) per metagenòmica de forma semblant a la comentada per als óssos bruns, seqüenciant els gens del 16 S rRNA a partir de mostres del cec de I. tridecemlineatus.
En aquest estudi els grups bacterians més abundants trobats, i que tenien més variació en l’estat d’hibernació respecte quan els animals eren actius foren Firmicutes, Bacteroidetes i Verrucomicrobia (com Akkermansia muciniphila, coneguda com a beneficiosa a la microbiota humana). Com veiem (Figura 5), en l’estació activa hi ha més Firmicutes mentre que amb la hibernació augmenten els altres dos grups. Com he comentat per als óssos, els Firmicutes prefereixen una dieta rica en polisacàrids mentre que els Bacteroidetes —i els Verrucomicrobia en aquest cas— consumeixen mucines produïdes per l’intestí. També hi ha menor diversitat de bacteris en hivernant. Les diferències de microbiota entre estació activa i hibernació són estadísticament significatives, molt més que les observades per l’edat, la dieta anterior a l’hibernació, i altres condicions. Tot plegat, la microbiota d’aquests suslics es reestructura cada any reflectint les diferències en les preferències dels microbis en quant als substrats de la dieta o de l’hoste i de les capacitats de supervivència dels diferents tàxons en l’ambient alterat de l’intestí en hibernació (Carey et al. 2013).
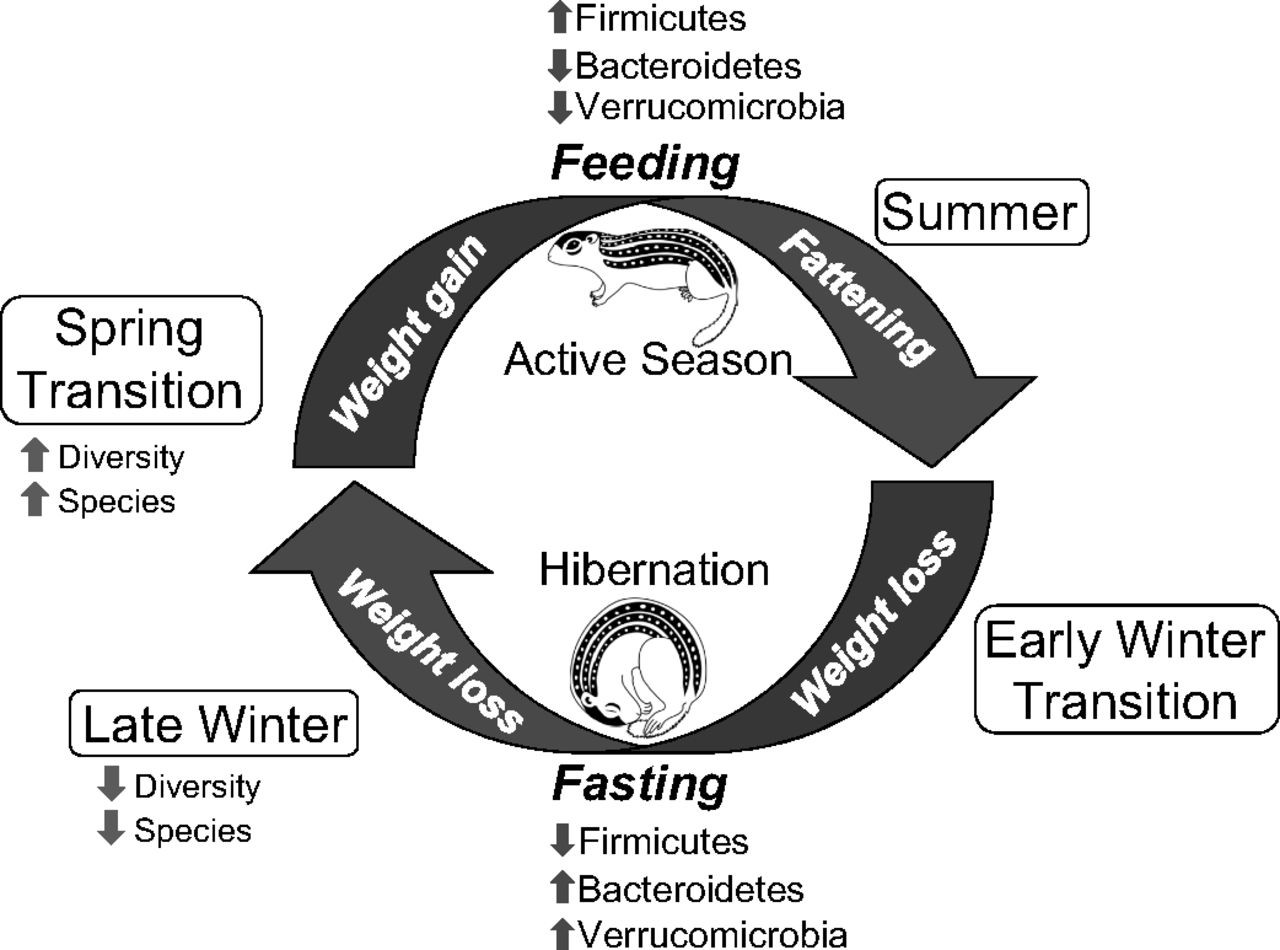
Figura 5. Esquema il·lustratiu dels principals canvis a la microbiota de l’esquirol de terra al llarg del cicle anual d’hibernació (Treta de Carey et al. 2013)
———————
LA MICROBIOTA DE LA HIBERNACIÓ PERMET LA RECUPERACIÓ DEL NITROGEN
Com hem vist abans, amb la hibernació manca el nitrogen de la dieta, i el mínim metabolisme de manteniment comporta un consum de les reserves de proteïnes —que pot portar a l’atròfia muscular—, i amb aquest consum s’alliberen compostos de nitrogen com l’amoni que és tòxic i el cos l’elimina en forma de urea per l’orina.
Però aquests animals no perden massa muscular durant l’hivern, i no excreten urea, ja que no orinen quan hibernen. Cóm s’explica ? Doncs la microbiota és la que permet la supervivència de l’animal durant la hibernació. Efectivament, s’ha vist que la microbiota dels comentats esquirols de terra de 13 bandes —Ictidomys tridecemlineatus— reciclen la urea produïda pel catabolisme de proteïnes, en aquest procés anomenat recuperació del nitrogen ureic (urea nitrogen salvage), on la ureasa microbiana té un paper clau.
De fet, aquest mecanisme ja era conegut als remugants, ja que la molt complexa microbiota del rumen inclou el reciclatge del nitrogen proteic dels mateixos microorganismes a través de la urea, i fins i tot aquest compost s’utilitza com a alternativa nitrogenada a la dieta dels remugants estabulats, donada la gran capacitat ureàsica de la rica i diversa microbiota remugant (Patra et al. 2018).
Però als animals monogàstrics com aquests esquirols no es coneixia aquesta capacitat de recuperació del N ureic, demostrada recentment per Regan et al. (2022). La urea resultant del catabolisme de les proteïnes de l’animal (Figura 6) és transformada per la ureasa dels microbis del cec intestinal en CO2 i amoni, i aquest és aprofitat per altres microbis per generar aminoàcids que són finalment transportats al fetge per sintetitzar proteïnes per als diferents òrgans.
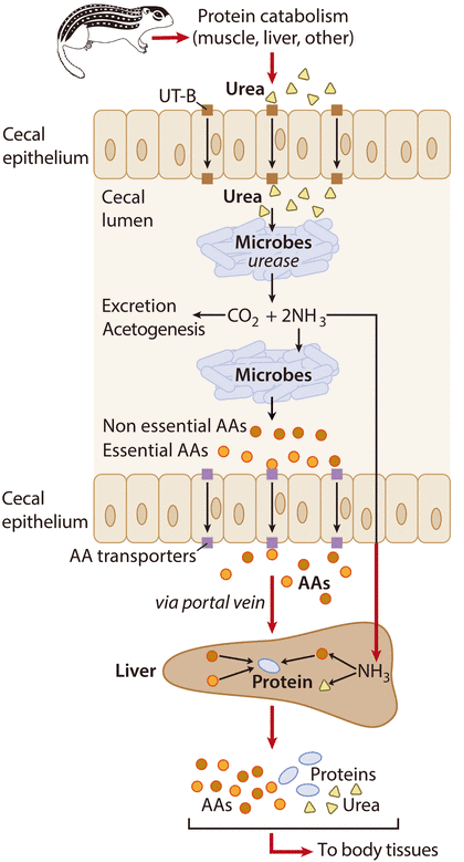
Figura 6. Mecanisme proposat de recuperació de N de la urea per la microbiota del cec del budell gruixut durant la hibernació del suslic Ictidomys tridecemlineatus (Figura treta de Regan et al. 2022).
Regan et al. (2022) també han vist que la recuperació del N ureic és més gran al final de la hibernació, just abans que l’esquirol es desperti i entri a l’estació activa, on li caldrà que els músculs s’hagin ben recuperat del torpor. Aquest augment de recirculació de N es reflecteix en la major abundància de transportadors d’urea i dels gens de la ureasa en aquest període (Figura 7). Els gens de la ureasa són 7: 1 transportador, 2 estructurals i 4 accessoris.
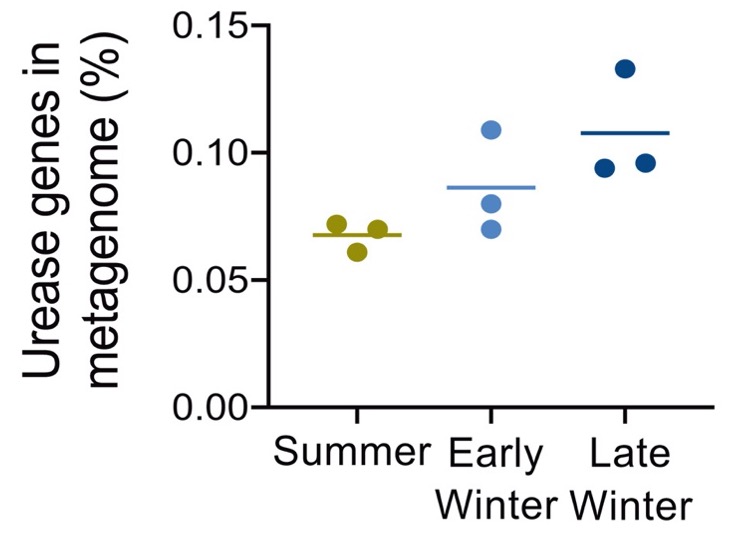
Figura 7. Proporció dels gens d’ureasa al metagenoma bacterià del budell de Ictidomys tridecemlineatus, a l’estiu, inici d’hivern i hivern tardà (Regan et al. 2022)
En analitzar la metagenòmica dels gens de la ureasa també s’ha vist que els tàxons bacterians més abundants són diferents en funció de quin d’aquests 3 períodes (Figura 8). Com veiem, el gènere Alistipes (un Bacteroidetes) és el que té més proporció de gens de la ureasa durant la hibernació, on també és el gènere més abundant, amb poblacions a l’hivern tardà sis cop superiors a les de l’estiu (Regan et al. 2022).
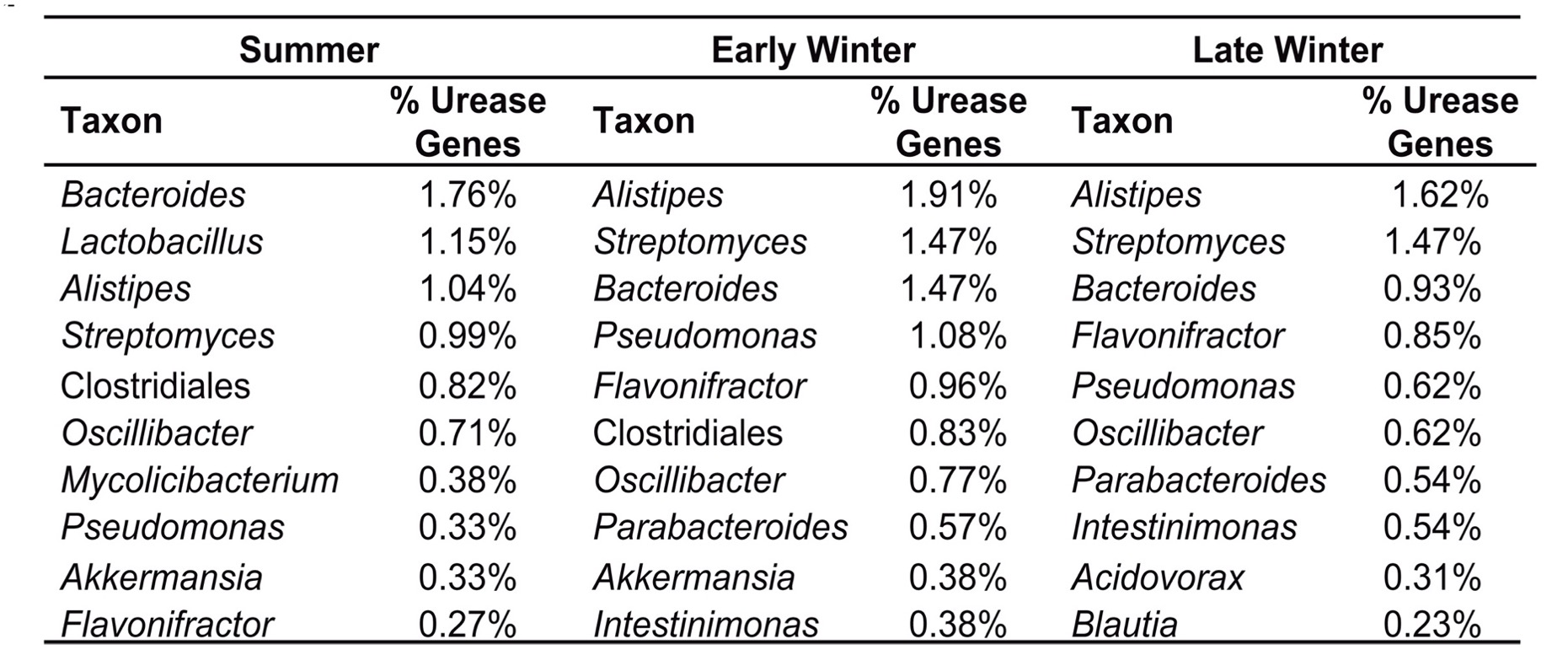
Figura 8. Els 10 tàxons bacterians més abundants en quant als gens d’ureasa a les seqüències del metagenoma bacterià del budell de Ictidomys tridecemlineatus, a l’estiu, inici d’hivern i hivern tardà (Regan et al. 2022)
A més a més, la recuperació del nitrogen de la urea pot facilitar l’estalvi d’aigua als esquirols hivernants desviant urea dels ronyons, amb la qual cosa cal menys aigua per a la producció d’orina, com també passa als camells quan els falta aigua (Mousa et al. 1983).
———————
I MÉS ENLLÀ DE LA HIBERNACIÓ
Aquest mecanisme de recuperació del nitrogen per la microbiota té implicacions que van més enllà de la hibernació. Per exemple, l’atròfia muscular afecta milions d’humans per causa de dietes limitades en nitrogen o també de sarcopènia a les persones d’edat avançada. Una millor comprensió dels mecanismes de com la microbiota de la hibernació mitiga els efectes de la limitació de nitrogen podria proporcionar estratègies per a la preservació dels músculs als humans (Regan et al. 2022).
D’altra banda, com hem vist, els óssos bruns —i altres animals— acumulen molt de greix abans d’hibernar i en canvi es mantenen metabòlicament sans. Entendre els mecanismes fisiològics subjacents i la possible relació amb la microbiota podria donar pistes per a noves teràpies de l’obesitat als humans (Sommer 2016).
I des d’un punt de vista més general, està clar que la coevolució dels mamífers —i altres animals— amb els microbis intestinals ha produït unes relacions ben complexes que donen beneficis a les dues parts simbionts. Els microbis modelen la biologia de llurs hostes de múltiples maneres, augmenten la resistència a la colonització per patògens, influeixen en la funció i estructura gastrointestinal, dirigeixen el desenvolupament del sistema immunitari, i augmenten la captació d’energia de la dieta. Al seu torn, els hostes animals proporcionen un ambient ric en nutrients que permet el desenvolupament de comunitats microbianes ben variades (Carey et al. 2013).
———————
BIBLIOGRAFIA
Carey HV, Andrews MT, Martin SL (2003) Mammalian Hibernation: Cellular and Molecular Responses to Depressed Metabolism and Low Temperature. Physiol Rev 83: 1153-1181.
Carey HV, Walters WA, Rob Knight R (2013) Seasonal restructuring of the ground squirrel gut microbiota over the annual hibernation cycle. Amer J Physiol-Reg It Comp Physiol 304, R33-R42
Garcia de Jesús E (2022) Gut microbes help some squirrels stay strong during hibernation. ScienceNews 27 Jan 2022.
Hibernation (2023, April 4). Wikipedia
Mousa HM, Ali KE, Hume ID (1983) Effects of water deprivation on urea metabolism in camels, desert sheep and desert goats fed dry desert grass. Comp Biochem Physiol A Comp Physiol 74:715-20
Patra AK, Aschenbach JR (2018) Ureases in the gastrointestinal tracts of ruminant and monogastric animals and their implication in urea-N/ammonia metabolism: A review. J Adv Res 13, 39-50.
Regan MD et al (2022) Nitrogen recycling via gut symbionts increases in ground squirrels over the hibernation season. Science 375, 6579, 460-463
Sommer F et al. (2016) The Gut Microbiota Modulates Energy Metabolism in the Hibernating Brown Bear Ursus arctos. Cell Reports 14, 1655-1661.
Thirteen-lined_ground_squirrel (2023, April 4) Wikipedia
Wilke C (2020) These Arctic squirrels recycle bits of their own bodies to survive winter. ScienceNews 16 Dec 2020.
